
This sample Biological Psychology Research Paper is published for educational and informational purposes only. If you need help writing your assignment, please use our research paper writing service and buy a paper on any topic at affordable price. Also check our tips on how to write a research paper, see the lists of psychology research paper topics, and browse research paper examples.
Ron, a married, 40-year-old schoolteacher, had a good life until it went bad. Nothing seemed to account for the overwhelming sexual urgency he began to experience—from searching Web sites for child pornography to soliciting prostitutes at massage parlors. Even though he knew that what he was doing was wrong, he lamely explained that the “pleasure principle overrode [his] restraint.” Ron’s wife found out about his aberrant behavior, had him evicted, and then prosecuted for pedophilia (fondling children). He was medicated and remanded to treatment for sex offenders. But he put himself at risk for a jail sentence when he violated the terms of treatment by soliciting sex from other program participants (Choi, 2002).
The night before sentencing, Ron went to an emergency room complaining of headaches and uncontrollable sexual urges. The attending physician referred him to the neurology division. There neurologists determined that he had a problem maintaining his balance and ordered an MRI scan. Further behavioral tests found that he couldn’t write or copy drawings and that he didn’t become particularly upset when he urinated on himself!
The MRI revealed an egg-sized brain tumor in the right side of Ron’s forebrain—specifically, in his orbitofrontal cortex. From the study of other patients with a damaged orbitofrontal cortex, the neurologists knew that poor impulse control, errors of judgment, and problems with social behavior were common outcomes. And because most people diagnosed with pedophilia experience their problems throughout a lifetime, yet Ron’s problems were recent ones, the neurologists confidently connected his behavioral disorder to the tumor.
Surgeons removed the tumor. However, seven months later, Ron’s headaches returned, and again he secretly began to collect pornography. Perhaps his problem was, as most of society would think, a moral failing, unrelated to how his brain functioned. But another MRI revealed another tumor, and another surgery to remove it successfully alleviated the symptoms (Choi, 2002). To the extent Ron had a moral problem, could it be attributed to a misplaced tumor?
A Biological Understanding Of Mind And Behavior
Like Ron, each of us has a personality—a unique culture-bound consciousness embedded in autobiographical memories. Because our mental life seldom requires conscious effort, we don’t even notice it. Throughout a lifetime we plan, think, fall in love, remember, forget, and become hungry, sleepy, angry, and sad. Hence, unless the biological underpinnings of human psychology are made the focus of study, as they are in this research-paper, we can live our lives quite successfully, thank you, without any knowledge of how our brains function.
We take for granted that consciousness, perception, motivation, memory, and the behaviors they make possible are dependent on an intact brain. We shouldn’t. The richness of the human experience begins in the brain’s functioning, and psychologists are not alone in wondering about the magic of its contents. Due to specific brain structures, humans see, hear, smell, taste, touch, and feel warmth, cold, pain, and pleasure. Through the process of introspection each of us with intact brains can become conscious of and ruminate on the complexity of our mental lives. We make moral judgments. Sitting on a mall bench we may also wonder about the minds of others. (Is that person by the department store who is talking to herself rehearsing a play, doing drugs, or suffering from a mental disorder? Should I be amused or concerned?)
One benefit of studying people like Ron is that a malfunctioning brain makes us aware of the normal brain’s invisible activities. Until the past few hundred years or so, we couldn’t weigh happiness or memories in drams, nor poke and prod at dreams using tools other than words. But now we can. Biological psychology is the study of the mind, brain, and behavior of humans and other animals from evolutionary, behavioral genetic, physiological, and psychological perspectives. In part, biological psychology attempts to understand mind and consciousness by studying its underlying “physical substance.” Neuroscientists study behavior in laboratory animals by measuring responses to brain manipulations using drugs, brain lesions, brain stimulation, and other intrusive techniques. Neuroscientists study humans as well as other animals. For example, the change in Ron’s behavior following removal of a tumor from his brain is an example of a brain-behavior relationship—an understanding of the mind and behavior from the perspective of its underlying physiological and anatomical substrates (Kolb & Whishaw, 2004). Cognitive neuroscientists study perception, attention, memory, language, and conceptual processes using brain imaging, often in conjunction with other behavioral measures such as paper-and-pencil tests. An example would be to study the neural correlates of human memory using a brain-imaging technique such as functional magnetic resonance imaging (fMRI). (The fMRI allows researchers to image neural activity of a person while in the act—for example, reading, listening to music, learning a task.) Although each theoretical perspective of psychology provides some unique insights not offered by others, cognitive neuroscience holds special promise for an integrative understanding of brain-behavior relationships. Studying the brain activity underlying appropriate social behavior, for example, allows us to understand better how tumors could produce Ron’s inappropriate social behavior.
This research-paper begins by describing, from evolutionary and genetic perspectives, the overall design of the human brain and the behavior it generates. We next explore the anatomical, physiological, and pharmacological properties of individual neurons comprising the brain, and how drugs affect the neurotransmitters secreted by neurons. Last, we will return to the brain structures and functions that underlay the conscious mind.
Evolutionary And Genetic Perspectives
All animals, including humans, have unique mental lives and behavioral tendencies due to the size, shape, and organizational features of their brains. Chimpanzees, for example, have mental lives and behavioral tendencies more similar to humans than to chickens. On the other hand, chickens also see, hear, learn, remember, get hungry and full, and respond to pain-inducing stimuli by squawking. It turns out that the uniquely designed brains of these three species also have similar brain structures.
Why is that? The answer to this question comes from scientists who study brain-behavior relationships from different perspectives: Geneticists compare genetic recipes for proteins that comprise brains in different species, neuroanatomists and neurophysiologists, respectively, study brain structure and function of animals, and neuroscientists study brain-behavior relationships. All are guided by Darwin’s evolutionary theory, which posits that brains were adapted to solve problems of survival and reproduction (Barkow, Cosmides, & Tooby, 1992; Darwin, 1859/1962). For example, an evolutionary answer to the question “Why is it human nature to sleep about 8 hours during the nighttime hours?” would be that humans who behaved in this way survived and reproduced more offspring than humans who didn’t. In other words, sleeping 8 hours (as opposed to 4 hours or 12 hours) is explained as an adaptive trait of human nature. Human brains and behavior differ from chimpanzee and chicken brains and behavior because they evolved in different ecological niches, including adaptively different sleeping patterns (as well as eating patterns, social behaviors, reproductive and maternal caretaking behaviors, and so forth). But their brains and behavior are also similar because problems of survival and reproduction have common elements.
What Is The Human Brain Plan?
Sixty years ago, C. Judson Herrick (1948) wrote The Brain of the Tiger Salamander, a much-forgotten book that detailed a brain plan common to all vertebrates (fish, amphibians, reptiles, birds, and mammals). Herrick pointed out that all vertebrate species have a central nervous system (CNS) comprising a bilateral (left and right side) brain and a spinal cord. All have 12 paired cranial nerves that bring sensory information into the brain and send motor (movement) signals to facial muscles. All are sensitive to different wavelengths of light from the sun. All have eyes and “see,” but not all “see” the same thing. All are sensitive to vibrations of molecules of moving air (that is, all have ears; a fish’s “ears” are sensitive to vibrations in water). All have taste and smell receptors; all are sensitive to touch, including warmth, cold, and pain. These receptors all enter their respective brains in the same place—the brainstem. Moreover, all have a forebrain, a midbrain, and a hindbrain (see Figure 13.1)—that is, a front end, middle, and back end. (Sensory information comes into the midbrain structures, and voluntary movement is initiated by the forebrain.)
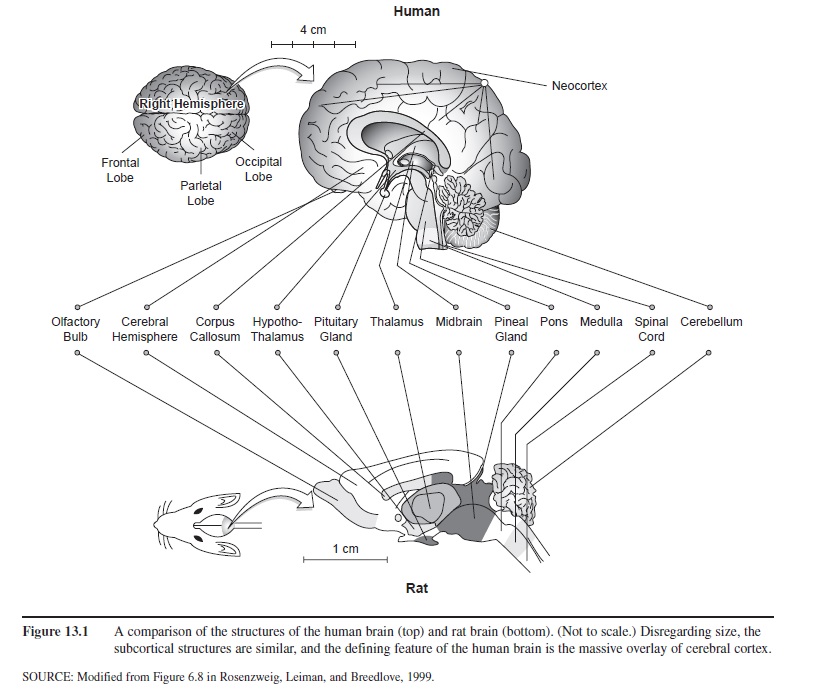 Figure 13.1 A comparison of the structures of the human brain (top) and rat brain (bottom). (Not to scale.) Disregarding size, the subcortical structures are similar, and the defining feature of the human brain is the massive overlay of cerebral cortex.
Figure 13.1 A comparison of the structures of the human brain (top) and rat brain (bottom). (Not to scale.) Disregarding size, the subcortical structures are similar, and the defining feature of the human brain is the massive overlay of cerebral cortex.
Humans also share with vertebrates the other mid-brain structures (commonly called the brain stem), the cerebellum, hypothalamus, and thalamus. Figure 13.1 shows placement of these common structures in the human brain (top) and the rat brain.
Given all that is common, what makes the human brain so uniquely different? The short answer is the huge amount of cerebral cortex of both hemispheres that completely covers the other brain structures. The human mind and behavior, then, are the result of an ancient brain plan common to other animals that has become overlain in our recent evolutionary history by hundreds of millions of “new” neurons (neocortex means “new cortex”) comprising the cerebral hemispheres. The uniqueness of human consciousness lies in the functioning of this massive overlay of neurons comprising the neocortex. To understand the neocortex better, we drop down to a lower level of analysis of brain functioning—the neurons of which the brain is made.
Neurons
Neurons are specialized cells in the brain that have abilities analogous to those of behaving animals. They are sensitive to the environment (that is, they receive information from their surroundings) and act on it. As shown on the left-hand side of Figure 13.2, each neuron has a soma, or cell body, and protrusions from it that either receive information (dendrites) or send information (the axon). One way that neurons are differentiated from most other cells in your body, then, is that a neuron is excitable—most appropriate for cells underlying psychological activity!
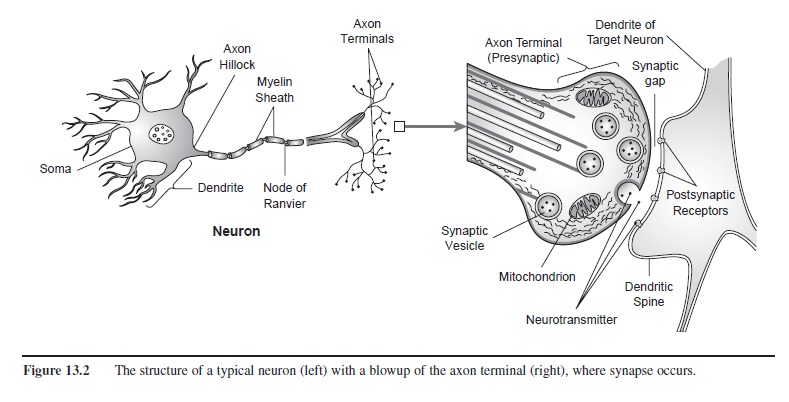 Figure 13.2 The structure of a typical neuron (left) with a blowup of the axon terminal (right), where synapse occurs.
Figure 13.2 The structure of a typical neuron (left) with a blowup of the axon terminal (right), where synapse occurs.
Depending on how neurons are analyzed, they reveal chemical, pharmacological, electrical, magnetic, and, ultimately, psychological properties. Electrical properties are measured with voltmeters. The electroencephalogram (EEG) measures the voltage of many thousands of neurons by attaching electrodes to the scalp. MRI, magnetic resonance Imaging, measures magnetic properties of thousands of neurons and by computer converts these potentials to images.
Though most function in similar ways, neurons are highly specialized. Different types of sensory neurons detect information in the environment (for example, the detection of light for seeing, vibration for hearing, and chemicals that are tasted and smelled). Motor neurons cause muscle tissue to contract, allowing us to move through the environment. (Perhaps because “moving a muscle” is conceptually simple, we tend to dismiss the role of neurons in accomplishing this basic behavioral unit. But that would be a mistake, for the abilities of skilled movements is in part what makes humans such interesting animals.)
Similarly functioning neurons in the brain are responsible for your ability to read this text, others for whether you are paying attention or nodding off. Some specialized neurons allow you to tie the first part of this sentence to the last part in what is called your short-term memory. Yet others are responsible for your impatience with this long sentence, in a long paragraph, in a long chapter. Hope to do well on your next assignment? You’ve got neurons responsible for your hope (or lack of it), for discipline (or lack of it), for perfecting your tennis swing, and for controlling your temper. Consciousness, perception, learning, memory, personality—all are the result of the functioning of billions of neurons in your central nervous system (Kandel, Schwartz, & Jessel, 2001).
Neurons that sense events function differently from motor neurons that activate muscles. Likewise, neurons in the cerebellum are not the same as those in the temporal lobe, and all differ from the five types of neurons in the retina of your eye. Instead of describing any one of these types, we’ll first describe the functioning of a generic, unspecialized neuron. When activated, it communicates with adjacent neurons by discharging a chemical, called a neurotransmitter. Some neurotransmitters cause a muscle to twitch. Others are involved in seeing or in memory formation. Still others are involved in the psychological experience of pleasure and pain. Then, we’ll examine how our knowledge of a neurotransmitter’s function is illuminated by different psychoactive drugs. We begin by learning about the neuron’s remarkable membrane “at rest,” before it becomes activated.
The Neuron’s Membrane: The Key To Excitability
Actually, the membrane of a neuron is never “at rest.” A neuron’s membrane separates its inside from its out-side. The membrane of a neuron has pores, or channels, through which ions can pass. Ions such as sodium and potassium are continuously attempting to move through this membrane covering dendrites, the soma, and the axon of a neuron. These ions, or electrolytes (so called because they carry a tiny positive or negative electric charge), are not “balanced” across the membrane. Their imbalance creates an electric charge across the membrane, called a membrane potential. Potassium and sodium ions are both positively charged, while chloride and proteins inside the cell are negatively charged. The charge, or resting membrane potential, across the cell membrane is due to this unequal concentration of electrolytes inside the cell (in the intracellular fluid) relative to outside the cell (in the extracellular fluid). If equal amounts of positive and negative charges existed both inside and outside the cell, the resting membrane potential would be zero. The normal resting membrane potential, or voltage across a neuron’s membrane before it is activated, however, is usually about -70 mV (minus 70 millevolts). (The charge is measured by sticking an electrode through the cell wall of the neuron and attaching it to a voltmeter.) The resting membrane potential is negative, not zero, because there are more negatively charged chemicals inside the cell and more positively charged ones outside. The cell is said to be at equilibrium (at rest) when there is no net movement of ions in or out of the cell.
However, at -70 mV, the neuron is “primed,” or poised to discharge. Then, one of two events can occur. A slight chemical disturbance of the membrane can cause a gradual change in the resting membrane potential of the dendrite or soma of a neuron. This effect is called a graded potential. The voltage may slowly increase or decrease, depending on the kind of chemical disturbance on the cell membrane. However, if the graded electrical charge is sufficiently large, the entire neuron can be triggered to discharge electrically. This large nerve impulse, sometimes referred to as the firing or spiking of the neuron, is called an action potential. An action potential, in turn, can ultimately disturb the resting membrane potential of other neurons, affecting their excitability. In fact, the action potential is one of the most important ways by which one neuron “talks” to another.
The Action Potential
Recall that the cell membrane of a neuron, including its axon, has pores, or channels, though which ions can pass. The rapid movement of ions through the axon’s membrane is the mechanism underlying an action potential. At rest, more sodium ions (Na+) are outside, and more potassium ions (K+) are inside the axon’s membrane, electrically polarizing it. Depolarization of the membrane to its threshold level has the effect of opening both sodium and potassium channels in the membrane for about 0.5 msec. During this brief time, the Na+ ions rush into the cell and the voltage across the membrane rises rapidly, from -70 mV, past zero volts, to about +50 mV. In response, seeking equilibrium, potassium ions (K+) rush out of the cell through their channels, eventually reversing the membrane’s polarity. This K+ ion movement in turn triggers the adjacent Na+ channels to close, rendering the membrane resistant to any further outflow of Na+ ions. In another fraction of a second, the resting membrane potential is restored, and the neuron is ready to discharge again. The result is a “spike” of electricity that flows rapidly away from the cell body down the axon.
When an action potential occurs, the depolarizing effect on the cell membrane quickly spreads to adjacent areas of the axon. The result is that the action potential travels down the length of the axon (the nerve “fires”). Shortly after the action potential has passed one portion of the axon, the K+ and Na+ ions resume their resting membrane condition. The axon cannot “half-discharge.” Once the neuron’s threshold for discharge has been reached, a complete action potential follows. In contrast to graded potentials, this all-or-nothing relationship is known as the all-or-none principle. Following an action potential, a brief time, called a refractory period, must pass before another action potential can be generated. Nevertheless, a given neuron can discharge repeatedly if the neuron remains depolarized below its threshold. Some neurons can fire up to 1,000 times per second.
Most axons are literally wrapped by a different type of cell (called a Schwann cell) that does not function like a neuron. Schwann cells form a myelin sheath that acts as an insulator, not allowing the electrical discharge of one neuron to affect adjacent axons. Figure 13.2 (left) shows that the myelin sheath covers the entire length of the axon except for tiny gaps between the Schwann cells, called the nodes of Ranvier. K+ and Na+ ion movement can occur only at the nodes of Ranvier. The result is that the action potential jumps rapidly from node to node, a process called saltatory conduction (saltatory from the Latin saltare, meaning “to leap”). The gain in speed is substantial: The one-meter-long, large diameter, myelinated axons that run from a 7-foot basketball player’s spinal cord to his toes conduct nerve impulses at about 100 meters per second. In contrast, small, unmyelinated fibers conduct impulses at a rate of only 1 to 2 meters per second. Such fibers are responsible for communicating chronic, dull aching pain.
The Release of Neurotransmitters
For present purposes we will assume that each neuron makes and releases but one type of neurotransmitter. The right-hand portion of Figure 13.2 indicates what happens when an action potential is conducted past the myelin sheath and arrives at an axon terminal. The axon terminal makes a synaptic connection with a target neuron or a muscle. (Synapse is the point at which one neuron encounters another.) Simply put, the action potential causes the release of a particular neurotransmitter into the synaptic gap (the space between adjacent neurons), effecting communication with another neuron. By contrast, motor neurons synapse on muscle fibers, causing them to contract.
How does this transmission of information from one neuron to the next occur? First, each neuron synthesizes its own particular neurotransmitter, which is stored in vesicles within the axon terminal. The action potential arriving at the axon terminal has the effect of opening calcium (Ca++) channels, allowing Ca++ to enter the cell, which causes the vesicles to fuse with the membrane wall of the axon terminal. The vesicle’s cell wall then opens, spilling its contents into the synaptic gap.
Should the frequency of action potentials arriving at the axon terminals increase, more Ca++ would enter the cell, causing more vesicles to merge with the terminal’s membrane and open, dumping more neurotransmitter into the synaptic gap. Once the neurotransmitter is in the synaptic gap, its distinctively shaped molecules find receptor sites in the target neuron’s membrane. Each neurotransmitter has a particular shape that can be thought of as a key—the postsynaptic membrane has receptor sites that can be characterized as locks into which the keys may or may not fit (Changeax, 1993).
How To Excite A Neuron
Disruption of the membrane potential of a target neuron and the target neuron’s generation of an action potential begins with the chemical events taking place at synapse. Figure 2 (right) shows how axon terminals synapse with both dendrites and the cell body of another neuron. When a neurotransmitter is released into the synaptic gap, it makes contact with the membrane of another neuron. The axon terminal is referred to as presynaptic (because it initiates events at synapse), and the target membrane on the other side of the synaptic gap is postsynaptic (because it is on the receiving end of synapse).
The contact that causes the target membrane to be excited is called an excitatory synapse. An excitatory synapse has the effect of decreasing, or depolarizing, the resting membrane potential of the target neuron. This depolarization is localized, meaning that it affects only a small area of the target neuron’s membrane. It results in a graded potential called an excitatory postsynaptic potential, or EPSP.
Meanwhile, a different neurotransmitter secreted from another neuron may be having the opposite effect when it makes contact with the target neuron’s membrane—it may inhibit the target neuron’s membrane potential. Like excitatory neurotransmitters, inhibitory neurotransmitters affect only a small area of the target neuron’s membrane. Inhibitory neurotransmitters tend to hyperpolarize, or increase the resting membrane potential. For this reason, hyperpolarization—an increase in the membrane potential—is called an inhibitory postsynaptic potential, or IPSP. Neurotransmitters that hyperpolarize the membrane, therefore, tend to work against, or to inhibit, the depolarizing effects of EPSP.
How do EPSPs and IPSPs interact? Think about EPSPs and IPSPs as sexual foreplay. Some events are excitatory, bringing you closer to achieving orgasm (the action potential); other events inhibit you. EPSPs and IPSPs are the localized, graded potentials that spread passively toward the axon hillock (see Figure 13.2), the site that determines each neuron’s threshold of excitability (approximately -65 mV, or 5 mV below the resting membrane potential). Once this threshold is met—once the EPSP has depolarized the membrane at the axon hillock to -65mV—the neuron initiates the voltage changes known as an action potential, as previously described.
The axon hillock integrates the effects of multiple inputs from other neurons through the processes of spatial and temporal summation. Spatial summation (summation on the surface space) occurs when neurotransmitters from several adjacent axon terminals excite an area of the membrane near the axon hillock. Temporal summation (summation in time) is caused by a rapidly firing neuron’s excitatory neurotransmitter depolarizing effects accumulating at the axon hillock. IPSPs, which act to block the excitatory effects on the axon hillock, can also summate spatially and temporally. The EPSPs and IPSPs algebraically summate: If the EPSP overcomes the IPSP at the axon hillock, the neuron generates an action potential.
Neurotransmitters And Their Effects
To describe in detail the chemical differences of the various neurotransmitters and how they interact with drugs (a field called neuropharmacology) is beyond the scope of this research-paper (see Cooper, Bloom, & Roth, 1996). Here we can provide only an overview: Table 13.1 groups some common neurotransmitters into five classes and describes their primary functions. Neurons that secrete these neurotransmitters differ both in their location in the brain and in their specific functioning.
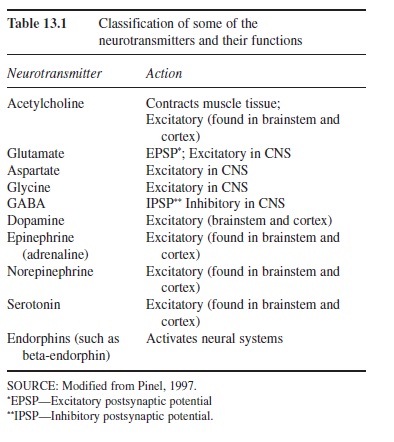 Table 13.1 Classification of some of the neurotransmitters and their functions
Table 13.1 Classification of some of the neurotransmitters and their functions

Let us look at a few examples. The neurotransmitter dopamine is excitatory and is synthesized primarily in the brainstem of humans and other animals. Dopamine pathways (from dopamine-secreting neurons) extend into the cerebral cortex. In experimental work with rats, neurons secreting dopamine can be stimulated by brief pulses of low-level electricity through an implanted electrode in a structure near the hypothalamus called the medial forebrain bundle (Olds, 1969; Olds & Milner, 1954). Electrical stimulation is presumed to mimic the action potentials of neurons. Rats will press a lever to receive such stimulation in the same way that they will press a lever to receive food. These and subsequent researchers studying dopamine pathways proposed that the neurotransmitter dopamine “causes” pleasure both in rats and humans—that “pleasure” has both an anatomical and a chemical nature.
To take another example, acetylcholine (ACh) is the neurotransmitter neurons secrete onto muscle fibers and is present in other neurons in the central nervous system and autonomic nervous system. Using the lock-and-key analogy, when released, dopamine fits dopamine receptors, and acetylcholine will activate only ACh receptors.
The actions of both dopamine and acetylcholine differ from those of yet another class of neurotransmitters, amino acids, which include glutamate (see Table 13.1). Glutamate has excitatory effects, and GABA, another amino acid, is an inhibitory neurotransmitter. Other excitatory neurotransmitters include epinephrine, norepinephrine, and serotonin. A distinctive class of chemicals called neuro-peptides also function as neurotransmitters. Endorphin, for example, is a neuropeptide that stimulates other neurons in the brain and spinal cord to produce analgesia (pain relief) and the experience of pleasure. Another neuropeptide, cholecystokinin, is released following a meal and produces satiety, the sensation of feeling full.
This brief summary of the neurotransmitters does not do justice to the incredible stories presently unfolding in neuroscience and neuropharmacology laboratories around the world. Sixty years ago, only two transmitters, one excitatory and one inhibitory, were known to exist. Table 1 lists just a few of many dozens of neurotransmitters that have since been discovered in humans and other animals.
Neurotransmitters made and released by neurons in discrete parts of the brain are the sources of our motivation and emotion, our voluntary behavior, the ways in which we learn and remember, and other aspects of our consciousness. Because humans share DNA with other animals and even plants, it should be no surprise that we also share some of the same neurotransmitters. For example, Hsieh, Lam, van de Loo, and Coruzzi (1998) reported finding genes in a weed, Arabidopsis, which encode for receptors sensitive to the neurotransmitter glutamate. Indeed, the number of plants that contain neurotransmitter-like chemicals that are common psychoactive drugs is amazing. Cannabis from marijuana and poppies that produce opium and tobacco leaves containing nicotine are well-known examples. Let us look at how some of these drugs affect the normal functioning of neurotransmitters.
Drugs, Neurons, And Behavior
Table 13.2 summarizes how drugs affect human consciousness through their action on neurons. Drug categories include depressants (natural and human-made), antianxiety drugs (human-made), stimulants (natural and human-made), antidepressants (human-made), narcotics (natural and human-made), and psychedelics and hallucinogens (natural and human-made).
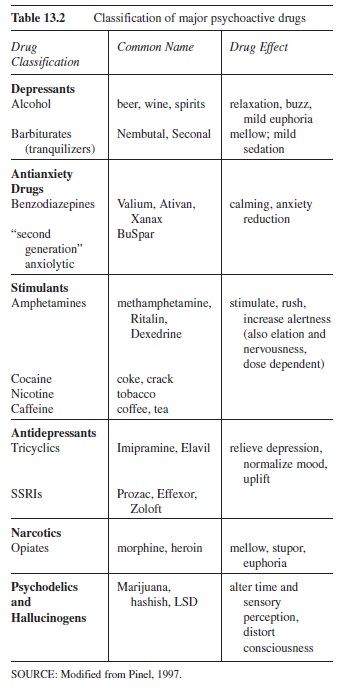 Table 13.2 Classification of major psychoactive drugs
Table 13.2 Classification of major psychoactive drugs
Depressants such as ethanol (beer, wine, and spirits) alter waking consciousness by depressing it, or slowing it down, by inducing relaxation and reducing physical activity. Alcohol stimulates the release of dopamine. Dopamine is also released by other drugs, such as cocaine, and, as described earlier, by electrical stimulation of the brain, by the ingestion of tasty food, or by reading a good book. Alcohol also alters consciousness by activating GABA receptors, depressing neural activity in the frontal cortex, which normally inhibits behavior. Thus, a moderate dose of alcohol disinhibits inhibited behavior; a person may therefore exhibit heightened sexuality, aggressiveness, or playfulness. These drug effects are dose-related; higher doses produce slower sensory-motor reaction times, disordered thought, slurred speech, poorer-than-normal judgment, and poor coordination. At the highest doses, alcohol becomes extremely dangerous: Sleepiness, unconsciousness, coma, and death can occur quickly as neurons in the brainstem that regulate consciousness are disabled.
Caffeine is a stimulant, a sympathomimetic (mimics the action of the sympathetic nervous system—the “fight or flight” system of arousal). The sympathetic division of the autonomic nervous system is aroused due to caffeine-stimulated release of the neurotransmitter epinephrine from adrenal glands. The result: dilation of the lungs’ bronchi, constricted blood flow to the brain, and enhanced blood flow to the rest of body. Another stimulant, amphetamine, overactivates dopaminergic and serotonergic neurons. Chronic use of high doses of amphetamine can cause permanent damage to the neurons that release dopamine and serotonin, producing “amphetamine psychosis”— severe paranoid hallucinations associated with aggressive, destructive behavior, as well as sleep disturbances, sexual dysfunction, depression, poor memory, loss of coordination, and even schizophrenia (Julien, 1995).
The primary effect of the stimulant cocaine is to block the reuptake of dopamine, allowing the neurotransmitter to remain longer in the reward and pleasure centers near the hypothalamus (see Figure 13.1). Nicotine produces its pleasurable effect by releasing epinephrine from various sites in the body. (Epinephrine, among other effects, causes CNS arousal.) It also releases an endogenous opiate, beta-endorphin. (Endogenous means “normally occurring.”) Because nicotine also stimulates acetylcholine receptors and dopamine receptors, tolerance to nicotine develops rapidly, making it one of the most highly addictive drugs.
Opiates (narcotics) such as morphine and heroin alter consciousness by producing analgesia and pain relief and, at higher doses, stupor and euphoria. These drugs’ “keys” perfectly fit the “locks” of opiate receptors, receptors that are also stimulated by naturally occurring “endogenous opiates” called endorphins. (The term endorphin comes from “endogenous morphine.”) Endorphins are released naturally when physiological systems are stressed. An example is endorphin release during and following an intense workout such as a competitive athletic event. The feeling of well-being during recovery is caused by endorphin release.
Hallucinogens such as marijuana, hashish, and LSD are all quite different compounds. Their chemical structure can be similar to serotonin, norepinephrine, and acetylcholine. How the various hallucinogens work is less well understood. For example, LSD and the designer drug ecstasy (X) are serotonin antagonists—they block serotonin receptors. The cannabis in marijuana fits perfectly in cannabinoid receptors found throughout the brain (the nucleus accumbens in the brainstem, cerebral cortex, hippocampus, cerebellum, basal ganglia, hypothalamus, brainstem, and spinal cord—even the spleen!). These cannabinoid receptors function as neuromodulators, altering the function of norepinephrine, dopamine, serotonin, acetylcholine, and other neurotransmitters.
From these examples one can appreciate how the biology of the brain provides a better understanding of mind and behavior. Normal brain functioning comprises myriad interactions of billions of interconnected neurons secreting functionally different neurotransmitters. Indeed, the model of EPSPs and IPSPs summating on a single neuron presented above does not begin to capture the complexities of brain functioning. Each neuron may have 10,000 or more places of synapse upon it, integrating information from thousands of different neurons.
One last example focuses on the neurotransmitter serotonin. Human-made drugs with the brand names Prozac, Zoloft, Welbutrin, and Effexor affect how much serotonin is in the synaptic gap between two neurons. In people who are neither too sad nor too happy, an “optimal” amount is released by serotonergic neurons. If too little serotonin is present, a person feels sad and depressed. By adding an SSRI, a selective-serotonin reuptake inhibitor (Prozac is an example), the amount of serotonin in the synaptic gap increases, and symptoms of depression are diminished.
Is Human Consciousness Merely Brain Chemistry?
That drugs interact with neurons to affect our conscious experiences raises other interesting questions. Are our mental and emotional lives merely chemical interactions? This question has not been (and may never be) answered by neuroscience researchers. In contrast to the underlying chemistry of consciousness, humans experience the phenomena of consciousness in what seem to be qualitatively different ways. The pain of a bee sting is so unlike the memory of last night’s dream, and our experience of each seems so unlike the chemical changes of neurons. Humphrey (1992) proposed that human consciousness is an emergent property of the interactions of billions of neurons in the large human brain. (Emergence is the unforeseen result of the synthesis of individual elements.) But not everyone thinks that brain chemicals provide a satisfying “explanation” of human (Chalmers, 2007) and nonhuman, animal consciousness (Wasserman, 1993). Certainly, “chemical, pharmacological, electrical, and magnetic” are forms of energy, but as yet, seeing, hearing, and tasting are not translatable into any of them. Therefore, it is reasonable to conclude that knowledge of the brain doesn’t “solve” the mind-body problem; rather, it is a highly promising approach.
The Human Brain
So far we have explored the evolutionarily and genetically determined brain plan of vertebrates, including humans. The overall design, including bilateral organization, spinal cord, brainstem, cerebellum, thalamus, hypothalamus, and cranial nerves, is common to all vertebrates. This early brain plan was adaptively selected by the environment in which life evolved—one that was sensitive to sunlight; vibrations in water (and later, in the earth’s atmosphere); chemicals (food and other animals) to be tasted and smelled; and motor systems to move, capture prey, avoid predators, find mates, and so forth. We have also examined the properties of the basic functioning units of these vertebrate brains, their neurons, and how drugs affect their neurotransmitters. Next we explore further how the human brain differs from the brains of mice, monkeys, and chimpanzees to afford us our unique consciousness and behavior.
Compare again the rat brain with the human brain in Figure 13.1. Note they are not drawn to scale. The mouse brain is much smaller than a pencil’s eraser. You can approximate the size of your own brain by putting your two closed fists together at the knuckles, each fist representing a left and right cerebral hemisphere. But the comparison is not fair, you might argue, because the human body is far larger than a rat’s is, so of course our brain is larger. You would be right in that there is a brain-mass/body-mass ratio that can be computed that would correct for body size. Even so, in such a ratio, humans remain “over-brained.” All the great apes (including humans, chimps, and gorillas) fare well in such brain-body ratio comparisons, but humans are the most outstanding (Jerison, 1973). Furthermore (and this is the key to human uniqueness), most “extra” neurons are in the outermost covering of the brain, the cerebral cortex. The subcortical (below the cortex) brain of a chimpanzee is extremely similar to that of a human. The chimpanzee brain is also covered by the neocortex (new cortex), which makes up the cerebral hemispheres. But for humans, the surface area of the neocortex, comprising billions of neurons, far exceeds that of a chimpanzee.
The wrinkled appearance of a human brain, with its hills (gyri) and valleys (sulci), is deceptively small. If you were to “skin” the cerebral cortex off the subcortical areas it completely covers, unfold it, and spread it out, it would be about the size of a large road map. (In demonstrating this point in lectures, I bring in a road map squashed into a baseball-sized ball, and unfold it.) This huge amount of “new” cortex developed fairly recently (0.5-1 mya) in our evolutionary history. It is this explosion of brain tissue that differentiates us from our hooting, cultureless, primate relatives. This neocortex and its interconnected-ness with an immediately underlying structure called the thalamus (see Figure 13.1) is what makes possible human consciousness, including thought, language, skilled movement, and culture (Edelman & Tononi, 2000). Why can’t a chimpanzee be taught language or sing a song? It doesn’t have a sufficient amount of neocortex or the appropriate organization of its limited neocortex.
This neocortex comprising the left and right cerebral hemispheres is conventionally divided into four lobes: frontal, parietal, temporal, and occipital (see Figure 13.1). Think of these “lobes” as rough anatomical and functional guidelines. For example, most of the occipital lobe (at the back of the brain) is made up of neurons involved in various activities of “seeing,” and neurons in a small portion of the left temporal lobe (on the left side of the brain at ear level) are necessary for “hearing and understanding language.” This approach emphasizes the brain’s specificity of function—each part of the brain is involved in a discrete functional activity.
The Brain’s Plasticity
Alternatively, the neocortex covering the human brain also exhibits what is called “plasticity.” The neocortex can acquire new functions through experience with the environment. To take an extreme example, if the so-called “language neurons” on the left side of the brain are damaged in a small child, the child no longer understands spoken language. Amazingly, with time, a commensurate area in the right temporal lobe is somehow induced to assume this function and the child can relearn language. (Unfortunately, this capability is mostly restricted to juvenile brains; very little language is relearn able if the damage occurs after puberty.)
Another example of the brain’s plasticity is that neurons “reorganize” themselves during a lifetime depending on what behavior a person engages in—a literal “use it or lose it” situation. Into music for a couple of years? Your brain organization will reflect this activity, and will change to reflect new activities in new environments such as playing chess for an extended number of years. A final example of the brain’s plasticity is called neurogenesis (Eriksson et al., 1998). New neurons appear in a part of the brain’s neocortex called the hippocampus when you encode large amounts of information into memory.
A human’s subcortical structures (common to humans, mice, and monkeys) reflect more specificity of function and less plasticity. Table 13.3 makes this point: The behaviors common to humans and other animals in the left column, though not all subcortically mediated, are more “hard-wired.” The reason is that the brain of an animal is first and foremost designed to keep it alive. The subcortical regions appeared millions of years before the cerebral hemispheres of the human brain evolved, and “plasticity of behavior” certainly doesn’t characterize how fish, frogs, and snakes live their lives!
This ancient brain design is reflected in how humans sense the world. The body’s skin senses begin with receptor neurons that transmit information about touch, pain, and temperature. Their axons come into the spinal cord, ascend along pathways through the midbrain, and synapse in the thalamus (see Figure 13.1). Taste and sound receptors send their axons into the medulla, a brainstem structure, where they synapse and ascend to the thalamus. Receptors in the eye are connected to nerves that project to the thalamus. The point is, sensing the world “begins” in the thalamus, a phylogenetically older brain structure, before being “completed” by neurons in the newer cerebral hemispheres.
 Table 13.3 Behaviors common to all animals (left) and unique to humans (right)
Table 13.3 Behaviors common to all animals (left) and unique to humans (right)
To appreciate better the connectivity between the old and new brain in humans, we’ll look more closely at how we see. Four specialized neurons in the retina of the eye are sensitive to light. In dim light, the rod receptors signal the presence or absence of light. The remaining three visual receptors, called cones, require a relatively high intensity light, and each is maximally sensitive to the wavelengths comprising the visible spectrum that are labeled red, yellow-green, and blue. When stimulated, these receptors synapse on bipolar cells, which in turn synapse on ganglion cells—both are also specialized retinal neurons. Their activity is modulated by two other types of neurons in the retina, called horizontal cells and amacrine cells. This 5-layer retina begins to organize the visual information, but “seeing” is accomplished further in the brain.
The axons of the ganglion cells leave the eye and form the optic nerve. Most of the fibers involved in the conscious experience of sight project to the thalamus. Other optic nerve fibers go to the brainstem, where they synapse on neurons that in turn reflexively control “mechanical” aspects of the eye such as dilation and constriction of the pupil (to regulate how much light enters the eye). Some optic nerve fibers project to the hypothalamus and are involved in regulating our circadian rhythms, including alertness, sleep, and metabolic functions (our body is warmer when light enters the eye and colder in the dark). Approximately 150,000 “non-seeing” optic nerve fibers project to another brainstem structure called the superior colliculus. The superior colliculus’s function is to turn our head reflexively, unconsciously, to direct our gaze. This portion of the brain allows us to drive our cars “unconsciously” by coordinating what we see with how we steer, paying little attention to the process.
Conscious sight is accomplished by millions of optic nerves that synapse first in the thalamus and then by new fibers from the thalamus to striate cortex, the primary visual sensory area located in the occipital lobe. Seeing can be interrupted at any point. For example, people with damaged portions of striate cortex have blind spots in their visual field, even if the retina and thalamus are intact.
What is seen? And why is 40 percent of a human’s neocortex—many hundreds of millions of neurons—devoted to “seeing”? Why can’t the millions of neurons of the subcortical areas get this job done? Answers to these questions have come from neuroscientists studying the connections of neurons in striate cortex to what is called extrastriate cortex, or secondary visual sensory areas, located in the occipital lobe, parietal lobe, and temporal lobe. The picture that emerges is that the secondary (and tertiary) areas of extrastriate cortex integrate information arriving from two separate retinas into a unified percept of color, form, and movement—our visual world.
But our language reflects that “seeing” is more than the percept of form and color. “I’ll believe it when I see it with my own two eyes!” “Oh, I see!” So, “seeing” is also psychologically believing and understanding. In his book, The Man Who Mistook His Wife for a Hat, neurologist and writer Oliver Sacks (1987) described a patient who suffered damage to one area of extrastriate cortex. He could see just fine, but he didn’t know and didn’t understand what his eyes were telling him. He could see his hat, and he could see his wife’s head, but he confused the two! He suffered a visual agnosia—seeing, but not understanding what he was seeing.
What Is The Function Of The Neocortex?
The lesson taught by such brain damage gives insight into the functions of massive numbers of interconnected neurons comprising the four lobes of the neocortex. Visual generated by thalamus and striate cortex are meaningless by themselves. Their meaning derives from integrating them with prior images remembered over a lifetime. A face is familiar or not depending on a memory of having seen the face before. Extrastriate cortical neurons encode such memories. A glass is only a glass if one “knows” what a glass should look like. It has functions as well as visual features. It has a “feel” when picked up, makes a clinking sound during toasts, and produces a distinctive “taste” when wine is drunk from it. If the wine is sour, it violates our expectations. Expectations are the neurons that integrate the information from visual, tactile, auditory, and taste neurons and compare it with what has been previously experienced.
Much of the parietal, temporal, and frontal lobes are comprised of neurons that have been “trained” through experience to integrate our separate sensory experiences into a unified consciousness. The neocortex accomplishes much more than integration, though. Humans are able to “read” emotion by simply glancing at a face. (Women are better at this task than men!) Such ability is mediated by neurons in the frontal and parietal lobes that integrate “emotional information” (fear, rage, love) generated by ancient subcortical areas of the brain with visual information of “seeing” a face. Cortical brain damage diminishes this ability (Bowers, Bauer, Coslett, & Heilman, 1985). The frontal lobes can also be surgically disconnected from subcortical areas by a procedure called a prefrontal lobotomy, disrupting the individual’s conscious experience of emotion.
Summary
The human brain can be thought of as the integration of an “old” brain with a “new” brain. The thin veneer of neocortex covering the more primitive structures of the human brain is what allows us, unique among animals, to be civilized. Both old and new brain parts comprise neurons that function to sense one another’s neurotransmitters as well as sense the outside world. Neurons also connect brain to muscle, allowing highly skilled voluntary movement. Specialized neurons in the neocortex receive lifelong training, a social process called acculturation. These cells’ integrative functioning allows each of us a unique personality comprising hopes, plans, and expectations.
Let us conclude this brief tour of the biological underpinning of our psychological lives by revisiting Ron, whose personality changed when a tumor disrupted the normal functioning of his orbitofrontal cortex (part of the frontal lobes located just above the orbit of the eye). The neurons there integrate information from the oldest parts of the brain—emotion associated with sociality and sexuality—and from all other parts of the brain. Ron’s tumor interfered with neurons that had afforded him impulse control and good judgment acquired through a lifetime. These “civilizing” neurons became dysfunctional; both old- and new-brain structures ran amuck, generating antisocial behavior. It is left for philosophers and theologians to ponder whether his behavior was “immoral.”
References:
- Barker, L. (2004). Psychology. Upper Saddle River, NJ: Pearson Custom Publishing.
- Barkow, J. H., Cosmides, L., & Tooby, J. (1992). The adapted mind: Evolutionary psychology and the generation of culture. New York: Oxford University Press.
- Bowers, D., Bauer, R. M., Coslett, H. B., & Heilman, K. M. (1985). Processing of face by patients with unilateral hemispheric lesions. I. Dissociations between judgments of facial affect and facial identity. Brain Cognition, 4, 258-272.
- Chalmers, D. (2007). The hard problem of consciousness. In M. Velman & S. Schneider (Eds.), The Blackwell companion to consciousness (pp. 225-235). Oxford: Blackwell.
- Changeax, J. P. (1993). Chemical signaling in the brain. Scientific American, 269, 58-62. Choi, C. (2002, October 21). Brain tumour causes uncontrollable paedophilia. NewScientist.com news service. Retrieved June 7, 2007, from http://www.newscientist.com/article/dn2943-brain-tumour-causes-uncontrollable-paedophilia.html
- Cooper, J. R., Bloom, F. E., & Roth, R. H. (1996). The biochemical basis of neuropharmacology. New York: Oxford University Press.
- Darwin, C. (1962). On the origin of species by means of natural selection. New York: Collier Books. (Original work published 1859)
- Edelman, G. M., & Tononi, G. (2000). A universe of consciousness: How matter becomes imagination. Boston: Basic Books.
- Eriksson, P. S., Perfilieva, E., Bjork-Eriksson, T., Alborn, A. M., Nordborg, C., Peterson, D. A., et al. (1998). Neurogenesis in the adult human hippocampus. Natural Medicine, 11, 1313-1317.
- Herrick, C. J. (1948). The brain of the tiger salamander. Chicago: University of Chicago Press.
- Hsieh, M. H., Lam, H. M., van de Loo, F. J., & Coruzzi, G. (1998). A PII-like protein in Arabidopsis: Putative role in nitrogen sensing. Proceedings of the National Academy of Sciences, 95(23), 13965-13970.
- Humphrey, N. (1992). A history of the mind: Evolution and the birth of consciousness. New York: HarperPerennial.
- Jerison, H. J. (1973). Evolution of the brain and intelligence. New York: Academic Press.
- Julien, R. M. (1995). A primer of drug action (7th ed.). New York: Freeman.
- Kandel, E. R., Schwartz, J. H., & Jessel, T. M. (2001). Principles of neural science (4th ed.). New York: McGraw-Hill.
- Kolb, B., & Whishaw, I. Q. (2004). Fundamentals of human neuropsychology (6th ed.). New York: Freeman.
- Olds, J. (1969). The central nervous system and the reinforcement of behavior. American Psychologist, 24, 114-118.
- Olds, J., & Milner, P. (1954). Positive reinforcement produced by electrical stimulation of septal area and other regions of the rat brain. Journal of Comparative & Physiological Psychology, 47, 419-427.
- Pinel, J. (1997) Biopsychology (4th ed.). Boston: Allyn & Bacon.
- Rosenzweig, M. R., Leiman, A. L., & Breedlove, S. M. (1999). Biological psychology. Sunderland, MA: Sinauer.
- Sacks, O. (1987). The man who mistook his wife for a hat. New York: Harper Perennial.
- Wasserman, E. A. (1993). Comparative cognition: Beginning the second century of the study of animal intelligence. Psychological Bulletin, 113, 211-228.
See also:
Free research papers are not written to satisfy your specific instructions. You can use our professional writing services to order a custom research paper on any topic and get your high quality paper at affordable price.
ORDER HIGH QUALITY CUSTOM PAPER

Always on-time

Plagiarism-Free

100% Confidentiality

{kind=link}